
发布时间:2025-10-29
在哺乳动物基因组中,基因表达受到启动子(promoter)和增强子(enhancer)等调控元件的精密调控。增强子作为一类关键的调控开关,其最显著特征在于能够跨越长距离激活靶基因的转录。目前普遍认为,此类远距离调控依赖于染色质在三维空间中的折叠,使增强子与启动子得以在物理上相互靠近并发生接触,而这些互作通常被限制在拓扑关联结构域(Topologically Associated Domains, TADs)内部。
然而,一个长期未解的核心科学问题是:增强子与启动子之间的互作是否遵循明确的位置规则?在某一TAD内部,增强子是否能够等同地激活该区域内任意位置的启动子,还是存在位置偏好性?换言之,基因组元件的线性排列如何具体调控基因表达活性,其背后的功能逻辑仍不清楚。尽管已有研究提供了一定线索,但在具有复杂三维构象的天然基因座中,启动子与增强子的相对位置如何精确调控转录强度,目前仍属未知。要回答这一问题,需系统性地改变增强子与启动子之间的相对位置,并对由此产生的功能效应进行精确定量分析。

2025年9月18日,荷兰癌症研究所Bas van Steensel在《science》发表题为“Functional maps of a genomic locus reveal confinement of an enhancer by its target gene”的研究论文。该研究开发了一种创新的高通量“报告基因跳跃”技术,通过在小鼠胚胎干细胞的Sox2基因座内数千个不同位置上随机插入报告基因,绘制出了首张超高分辨率的增强子激活功能图谱。研究发现,Sox2基因本身会强烈地“限制”其远端增强子的作用范围,将其活性牢牢地约束在基因与增强子之间的区域内。一旦Sox2基因被删除,这种限制效应就会被打破,增强子的激活能力会显著增强并扩散至更广阔的基因组区域。进一步的机制研究表明,Sox2基因的编码序列(CDS)在介导这种限制效应中起关键作用,它能帮助基因更有效地与增强子互作,从而在“竞争”中胜出,阻止增强子激活其他启动子。
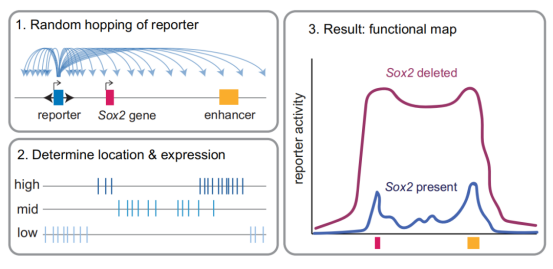
01、高分辨率功能图谱揭示:增强子激活效应严格受限,与三维互作频率相关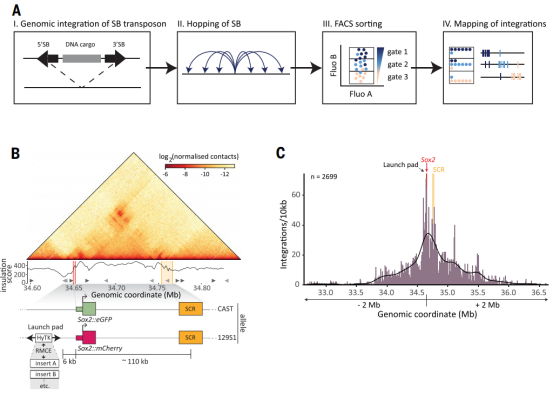
研究首先通过“Sleeping Beauty hopping”技术,将含Sox2启动子的报告基因随机插入基因座内数千个位点。团队采用基于Tn5转座酶的高通量测序,将每个细胞亚群中的整合位点与表达活性直接关联,在1.5 Mb区域内以前所未有的分辨率绘制了增强子激活功能图谱。
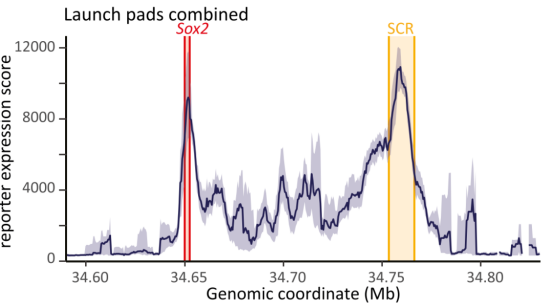
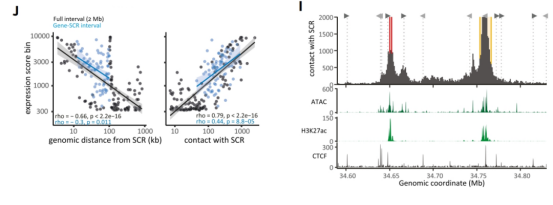
结果显示,增强子活性并非均匀分布,也不随距离简单衰减,而是呈现两个明显活性高峰:一个位于远端增强子核心区,另一个位于内源Sox2基因附近。通过区域捕获Micro-C测序分析三维染色质互作频率,发现该活性模式与预先存在的三维互作频率高度吻合,报告基因活性与其所在位置和增强子的互作频率成正比,表明物理接触频率是激活强度的关键决定因素。
02、Sox2基因对增强子的作用范围施加了强烈的限制
研究最核心的发现是,Sox2基因本身作为“限制者”,严格控制其增强子的作用范围。通过CRISPR/Cas9敲除内源Sox2基因后,整个基因座内报告基因活性普遍大幅提升,增强子影响范围显著扩大,甚至激活了正常情况下完全沉默的远端区域。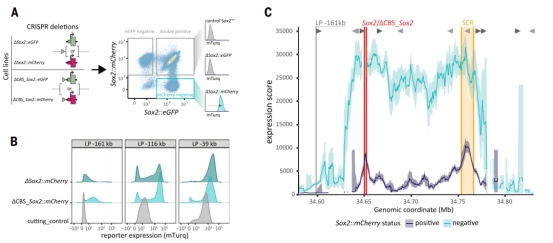
这一结果证明,内源Sox2基因形成了一种“门控效应”,主动将增强子功能限制在局部范围内,防止其对其他潜在靶点的“泄露”。进一步通过RCMC测序发现,Sox2缺失引起染色质三维结构重塑,增强子的接触范围向上游扩散,部分解释了其激活范围的扩大。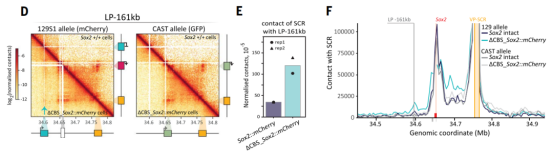
03、Sox2编码序列是介导增强子竞争优势的关键元件
为了探究Sox2基因实现自我限制的分子机制,研究团队对基因的不同组成部分进行了系统性剖析。他们发现,Sox2基因的启动子本身并不足以解释其强大的限制能力。其竞争优势的关键其实在于基因的内部。研究表明,仅在报告基因中加入约1 kb的Sox2的CDS,就能显著提升其表达活性,并使其有能力与内源的Sox2基因进行竞争,从而导致内源Sox2的表达水平下降。
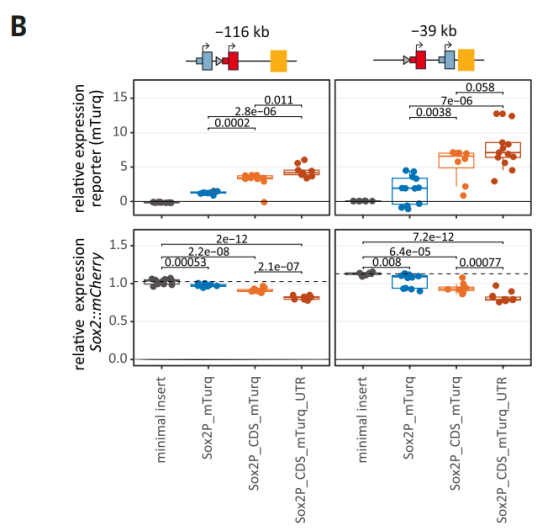
进一步的结构改造实验发现,CDS的这种增强效应与其转录方向、是否被翻译或在转录本中的位置无关,这表明CDS本身可能包含一个内在的、类似增强子或促进子(facilitator)的调控元件。
RCMC分析也初步证实,含有CDS的报告基因与SCR的接触频率有小幅增加,这可能部分解释了其增强效应。该元件的存在,使得天然Sox2基因能够更有效地与增强子SCR互作,从而在这场调控资源的竞争中占据主导地位。
04基因-增强子互作遵循“胜者为王”的竞争模型
综上,研究提出了增强子-启动子互作的动态竞争模型。增强子并非被动激活最近启动子,而是主动与区域内所有启动子互作。内源Sox2基因凭借其CDS这一“竞争利器”,能更高效“捕获”增强子,主导其活性,并抑制其对周围其他启动子的激活。一旦该最强竞争者被移除,被“解放”的增强子便可更自由地作用于竞争力较弱的启动子,重塑整个基因座的转录活性景观。这一发现揭示了基因本身在调控其与增强子互作中的主动作用。
这项研究为基因调控领域带来双重启发。在技术上,“报告基因跳跃”方法为绘制其他复杂基因组位点的功能图谱提供了强大、普适的工具,可精准剖析与发育、疾病相关关键基因的调控网络。在概念上,“基因自我限制增强子”模型更新了以往对增强子-启动子互作的认知,提示基因编码区本身可作为重要DNA调控元件。未来需重新审视基因内部序列在调控三维结构和表达活性中的作用。
此外,这一发现为基因编辑和合成生物学开辟了新方向:通过精确设计或修饰编码序列,未来或可实现对增强子活性的“重定向”,从而增强治疗性基因表达或抑制致病基因活性,为精准基因调控疗法提供新策略。例如,在设计CAR-T或基因替代疗法的表达盒时,可考虑整合这类功能性CDS元件,以提升治疗基因的表达效率与稳定性,显著增强疗效。
*转载自锐竞平台
